Berge und Täler im Heterochromatin
Max-Planck-Wissenschaftler entschlüsseln Mechanismen zur Bildung von Heterochromatin
Damit das zwei Meter lange DNA-Molekül in dem nur wenige Tausendstel Millimeter großen Zellkern untergebracht werden kann, müssen lange Abschnitte sehr eng gepackt werden. Epigenetische Markierungen halten diese als Heterochromatin bezeichneten Abschnitte aufrecht. Wissenschaftler des Max-Planck-Instituts für Immunbiologie und Epigenetik in Freiburg haben nun zwei weitere Mechanismen aufgedeckt, die für die Bildung von Heterochromatin notwendig sind. Die Forschungsgruppe um Thomas Jenuwein beschreibt die beiden Enzyme Prdm3 und Prdm16, die bestimmte Verpackungsproteine der DNA mit einer Methylgruppe versehen. Diese epigenetische Markierung sorgt dafür, dass das Heterochromatin und mit ihm die Struktur des Zellkerns erhalten bleiben. In einer weiteren Studie haben sie zudem festgestellt, dass Transkriptionsfaktoren im Heterochromatin binden und die Herstellung von nicht-kodierenden RNAs unterdrücken. Im Gegensatz zu weniger dicht gepackten Regionen, dem sogenannten Euchromatin, in denen sich Transkriptionsfaktoren an bestimmten Stellen häufen, sind die Bindungsstellen von Transkriptionsfaktoren im Heterochromatin wesentlich zufälliger verteilt.
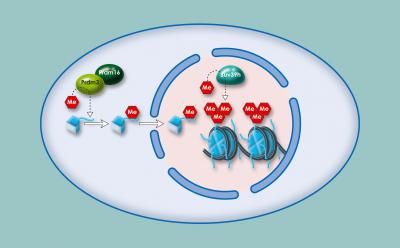
Histon-Methylierung im Heterochromatin: Die Enzyme Prdm3 und Prdm16 versehen das Histon H3 mit einer Methyl-Gruppe. Im Zellkern werden dem Histon zwei weitere Methylreste von Suv39h angehängt. Nur mit dieser dreifachen Methylierung bleibt das Heterochromatin stabil.
Art For Science
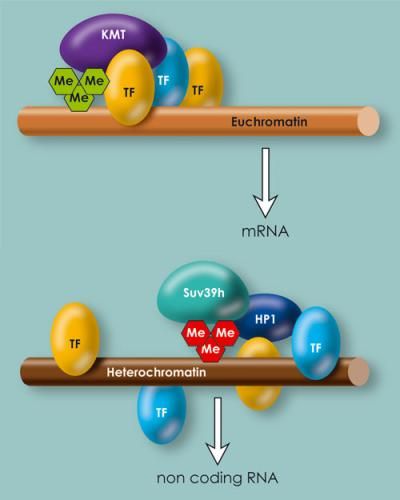
Im Euchromatin und Heterochromatin sind die Bindungsstellen für Transkriptionsfaktoren unterschiedliche verteilt. Im Euchromatin häufen sie sich an bestimmten Stellen, im Heterochromatin sind sie zufälliger verteilt.
Art For Science
Das Chromatin besteht aus dem DNA-Molekül und einer Vielzahl von Proteinen, darunter die als Verpackungsproteine fungierenden Histone. Im Gegensatz zum leicht zugänglichen Euchromatin, das den Großteil der Gene beinhaltet, finden sich im dicht gepackten Heterochromatin hauptsächlich repetitive Sequenzen, die nicht-kodierende RNA-Moleküle bilden können. Heterochromatin-Abschnitte liegen beispielsweise rund um die Zentromere und an den Chromosom-Enden, den Telomeren. Chemische Modifikationen an den Histonen können den Verpackungsgrad des Chromatins verändern. Dabei versehen z.B. sogenannte Methyltransferasen die Proteine an unterschiedlichen Stellen mit Methyl-Gruppen. Diese epigenetischen Veränderungen regulieren die Bildung und Aufrechterhaltung des Heterochromatins.
Ines Pinheiro, eine Doktorandin in der Abteilung von Thomas Jenuwein, hat nun entdeckt, dass Prdm3 und Prdm16 als Methyltransferasen wirken und an das Histon H3 in der Position Lysin 9 (H3K9) eine Methylgruppe anheften. Die beiden Proteine waren bislang nur als Transkriptionsfaktoren bekannt, die die Aktivität verschiedener Gene regulieren. Wie wichtig Prdm3 und Prdm16 sind, zeigen Experimente, in denen die Freiburger Forscher beide Enzyme ausschalteten: Das Heterochromatin bricht danach zusammen und die heterochromatischen Regionen können abgelesen werden. „Unsere Experimente zeigen, dass Prdm3 und Prdm16 eine Methyl-Gruppe an H3K9 heften. Dieses einfach methylierte H3 (H3K9me1) wird dann in den Zellkern transportiert und in das Heterochromatin eingebaut. Nur dann bleibt das Heterochromatin bestehen", erklärt Thomas Jenuwein, Direktor am Max-Planck-Institut für Immunbiologie und Epigenetik. An das einfach methylierte Histon können dann andere Methyltransferasen wie Suv39h noch zwei weitere Methyl-Reste (H3K9me3) anfügen und das Heterochromatin so weiter stabilisieren.
Die Freiburger Forscher beobachteten zudem, dass ohne Prdm3 und Prdm16 die Lamina des Zellkerns zerstört wird. Das Heterochromatin muss mit dieser Schicht aus Lamina-Proteinen unterhalb der Kernhülle verbunden sein. „Für ein stabiles Heterochromatin benötigt die Zelle offenbar die Methylierung an H3K9 und eines noch unbekannten Chromatin- oder Lamina-Proteins durch Prdm3 und Prdm16. Wir gehen davon aus, dass die beiden Enzyme wie andere Methyltransferasen nicht nur Histone methylieren können. Noch wissen wir allerdings nicht, ob die Zerstörung der Lamina durch den Verlust des Heterochromatins oder durch die fehlende Methylierung eines Lamina-Proteins ausgelöst wird", sagt Jenuwein.
Aber nicht nur die Methylierung von Histonen ist für die Aufrechterhaltung von Heterochromatin-Regionen erforderlich. In einer weiteren Studie haben die Doktorandinnen Aydan Karslioglu und Valentina Perrera die Rolle von Transkriptionsfaktoren untersucht, also Proteinen, die an die DNA binden und die Aktivität von Genen kontrollieren – im Falle des Heterochromatins die Unterdrückung von nicht-kodierenden RNA-Molekülen. Zwei davon sind demzufolge für ein intaktes Heterochromatin unverzichtbar: Pax3 und Pax9. Nur wenn diese beiden Transkriptionsfaktoren und ihre Bindungsstellen in der repetitiven DNA vorhanden sind, bleibt das Heterochromatin bestehen. Die Forscher gehen jedoch davon aus, dass auch noch weitere Transkriptionsfaktoren an repetitive Sequenzen im Heterochromatin binden können.
Transkriptionsfaktoren kontrollieren also die Genaktivität sowohl im Euchromatin als auch im Heterochromatin. Trotzdem gibt es Unterschiede zwischen den beiden: Im Heterochromatin sind die Bindungsstellen relativ zufällig über den DNA-Strang verteilt, im Euchromatin dagegen häufen sie sich an den für die Genregulation wichtigen Stellen. „In unseren Daten sieht die Verteilung im Heterochromatin etwa so aus wie die Aiguilles Droites im Mont Blanc-Massiv: Viele kleine Gipfel ohne tiefe Täler dazwischen. Das Euchromatin gleicht eher dem Matterhorn: Ein hoher Gipfel ohne Nebengipfel", beschreibt Thomas Jenuwein die Ergebnisse.
Ein wichtiger Unterschied zwischen Heterochromatin und Euchromatin liegt für die Forscher in der Kontrolle der Genaktivität und der Bildung von RNAs. „Im Heterochromatin sind die Bindungsstellen für Transkriptionsfaktoren eher zufällig verteilt, so dass sie sich nicht gegenseitig verstärken können. Die DNA kann deshalb dort nicht so abgestimmt abgelesen werden. Insgesamt überwiegen hemmende Einflüsse, die das Heterochromatin weitgehend abschalten", sagt Jenuwein. Im Euchromatin dagegen binden die Transkriptionsfaktoren so an die DNA, dass sie sich ergänzen. Dies erlaubt eine fein abgestimmte Kontrolle der Gen-Aktivität.
Originalveröffentlichung
I Pinheiro, R Margueron, N Shukeir, M Eisold, C Fritzsch, F M Richter, G Mittler, C Genoud, S Goyama, M Kurokawa, J Son, D Reinberg, M Lachner & T Jenuwein; "Prdm3 and Prdm16 are H3K9me1 Methyltransferases Required for Mammalian Heterochromatin Integrity"; Cell 2012 (Vol. 150, Issue 5, pp. 948-960)
A Bulut-Karslioglu, V Perrera, M Scaranaro, I A de la Rosa-Velazquez, S van de Nobelen, N Shukeir, J Popow, B Gerle, S Opravil, M Pagani, et al.; "A transcription factor–based mechanism for mouse heterochromatin formation"; Nature Structural & Molecular Biology (2012)

Weitere News aus dem Ressort Wissenschaft




















































