Durchgang nur für Auserwählte
Forscher entschlüsseln Frachterkennungs-Mechanismus eines molekularen Shuttles, das neben zelleigenen Komponenten auch das HIV-Genom transportiert
Massiver Güterverkehr herrscht nicht nur auf unseren Straßen. Auch im Inneren lebender Zellen wird ständig Fracht von einem Ort zum anderen transportiert. Die Kommandozentrale der Zelle - der Zellkern - ist dabei streng bewacht. Winzige Poren in der Zellkern-Hülle fungieren als hochselektive Tore: Während kleinere Moleküle meist ungehindert passieren können, sind größere auf ein Shuttle angewiesen - ein Service, den das Humane Immundefizienz-Virus (HIV) für seine Vermehrung in der Wirtszelle missbraucht. Forscher des Max-Planck-Instituts für Biophysikalische Chemie, der Universität Göttingen und der Technischen Universität München haben jetzt den entscheidenden Mechanismus aufgeklärt, wie der molekulare Transporter CRM1 nicht nur eine immense Vielzahl zelleigener Komponenten, sondern auch das HIV-Genom erkennt. Die Erkenntnisse könnten zukünftig neue Therapiewege zur Behandlung von AIDS eröffnen.
Komplex gebaute Zellen von Pilzen, Pflanzen und Tieren gleichen Miniatur-Hightech-Fabrikanlagen mit räumlich getrennten Montagehallen, Kraftwerken, Verpackungs- und Sortierstationen. Das Erfolgsrezept Arbeitsteilung hat allerdings seinen Preis: Eine ausgeklügelte Logistik ist nötig, um die Kommunikation und den Stoffaustausch zwischen den einzelnen Abteilungen einer Zelle (den Kompartimenten) sicherzustellen. Der Frachtverkehr zwischen den Kompartimenten steht dabei der Rushhour einer Großstadt in nichts nach. Allein der Stoffaustausch zwischen dem Zellkern und dem Zytoplasma ist gewaltig. So produzieren die Proteinfabriken (Ribosomen) außerhalb des Zellkerns, wodurch sämtliche für den Zellkern bestimmte Proteine aus dem Zytoplasma importiert werden müssen. Der Zellkern wiederum archiviert die Baupläne der Proteine in Form der Gene. Abschriften dieser Baupläne (Boten-Ribonukleinsäuren) müssen daher aufwendig zu den Proteinfabriken in das Zytoplasma exportiert werden. Nicht zuletzt liefert der Kern auch die Ribosomen-Untereinheiten selbst, die im Zytoplasma zu funktionstüchtigen Proteinfabriken zusammengesetzt werden.
Stoßverkehr nonstop an der Kernpore
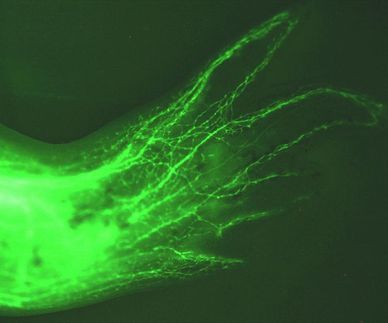
Was in den Zellkern hinein und hinaus darf, wird streng kontrolliert. Der Zellkern ist für Proteine und andere Makromoleküle durch zwei Membranen hermetisch abgeriegelt. Durchlass erlauben nur die etwa 3000 Kernporen, die in die Kernmembran eingebettet sind. Mehr als eine Million Zell-Bausteine werden in jeder Minute durch die Kernporen einer menschlichen Zelle hindurchgeschleust. Kernporen fungieren dabei als hochselektive Sortiermaschinen. "Jede von ihnen kann bis zu 1000 Frachtkomplexe pro Sekunde 'abfertigen'", erklärt Zellbiologe Dirk Görlich, Leiter der Abteilung Zelluläre Logistik am Max-Planck-Institut für biophysikalische Chemie. Während Kernporen die meisten kleinen Moleküle ungehindert passieren lassen, sind große Makromoleküle für ihren Transport durch die Kernpore auf Shuttle-Moleküle angewiesen. Diese haben das Privileg, die Kernporen nahezu ungehindert passieren und dabei Fracht mitnehmen zu können. Allerdings darf nicht jeder Passagier an Bord, sondern die Shuttles erkennen mit molekularer Präzision, welche Moleküle in den Kern, welche aus dem Kern und welche gar nicht transportiert werden sollen.
Viren mit selbst ausgestelltem Passierschein
Ein wahres Multitalent unter den Shuttle-Molekülen ist der Transporter CRM1. Er exportiert hunderte, wenn nicht gar tausende verschiedene Zell-Bausteine aus dem Kern in das Zytoplasma. Das Spektrum seiner Fracht reicht von einzelnen Proteinen bis hin zu den gigantischen Ribosomen-Untereinheiten für die Proteinfabriken. Das Humane Immundefizienz-Virus (HIV) missbraucht dieses Shuttle-Molekül allerdings für eigene Zwecke. Um sich zu vermehren, schleust das HIV sein Erbgut in das Genom der Wirtszelle ein und programmiert diese auf die Produktion von Virus-Partikeln um. Mithilfe zelleigener Enzyme wird das virale Erbgut in Ribonukleinsäure kopiert, über den CRM1-Transporter in das Zytoplasma der Wirtszelle exportiert und schließlich in neue Viruspartikel verpackt.
Molekulares Adress-Etikett
CRM1 erkennt sein Frachtgut anhand sogenannter Exportsignale, die einen Sequenzabschnitt von neun bis 14 Aminosäuren umfassen. Diese Signale sind allerdings derart variabel, dass sich die Frage stellt: Wie kann CRM1 derart variable Signale erkennen und somit Frachtmoleküle korrekt von allen anderen Objekten unterscheiden? Bei der Kontrolle seines Frachtguts lässt sich CRM1 nicht ohne Weiteres beobachten - die Transportkomplexe sind selbst für beste Lichtmikroskope viel zu klein. "Erst mithilfe der Röntgenstrukturanalyse konnten wir CRM1 mit seinem Frachtgut in atomarem Detail untersuchen. Wir haben so das Transportgut gewissermaßen durch die 'Brille' des Rezeptors betrachtet", erzählt Thomas Güttler. Dem Nachwuchswissenschaftler aus der Abteilung Zelluläre Logistik gelang es in aufwendigen Experimenten, CRM1 zusammen mit zwei verschiedenen Exportsignalen zu kristallisieren, die zum einen von zelleigener Fracht und zum anderen vom HIV stammen. Diese beiden Exportsignale folgen stark unterschiedlichen Mustern. "Überraschenderweise werden sie aber von einer einzigen 'Ladefläche' (Bindungsstelle) an CRM1 erkannt. Diese bleibt starr und unverändert, während sich die unterschiedlichen Exportsignale des Frachtguts an die Bindungsstelle des CRM1-Transporters anpassen", so Piotr Neumann, Wissenschaftler in der Abteilung Molekulare Strukturbiologie von Ralf Ficner am Institut für Mikobiologie und Genetik der Universität Göttingen.
Frachtgut oder nicht? Expressversand oder Standard? Das hängt davon ab, wie gut das Exportsignal der Fracht zur "Ladefläche" des CRM1-Shuttles passt. Je stärker dieses bindet, desto höher ist die Priorität, mit der die Fracht exportiert wird. "Wir kennen nun nicht nur die Regeln, nach denen der Shuttle eine astronomische Zahl unterschiedlicher Frachtgüter erkennen kann, sondern auch, wie er seine Prioritäten setzt", sagt Görlich. "Zu verstehen, wie ein Virus die zelluläre Infrastruktur für seine Vermehrung innerhalb der Wirtszelle missbraucht, ist auch von entscheidender Bedeutung, um zukünftig neue Therapien zur Behandlung von AIDS entwickeln zu können".
Originalveröffentlichung Thomas Güttler, Tobias Madl, Piotr Neumann, Danilo Deichsel, Lorenzo Corsini, Thomas Monecke, Ralf Ficner, Michael Sattler, Dirk Görlich; "NES consensus redefined by structures of PKI-type and REV-type nuclear export signals bound to CRM1"; Nature Structural & Molecular Biology, Online-Publikation, 24.10.2010
Weitere News aus dem Ressort Wissenschaft



















Diese Produkte könnten Sie interessieren

Antibody Stabilizer von CANDOR Bioscience
Protein- und Antikörperstabilisierung leicht gemacht
Langzeitlagerung ohne Einfrieren – Einfache Anwendung, zuverlässiger Schutz

DynaPro NanoStar II von Wyatt Technology
NanoStar II: DLS und SLS mit Touch-Bedienung
Größe, Partikelkonzentration und mehr für Proteine, Viren und andere Biomoleküle






























